Lycopene is one of the most well-known carotenoids in nature due to its high antioxidant properties and wide health benefits. Due to a wide range of applications in food, nutraceuticals, cosmetics, and pharmaceuticals, demand for lycopene has grown significantly and its unmet need requires production at large scale. Conventionally, lycopene extraction from natural sources suffers from extensive downstream processing and poor yield. Industrial processes using tomato pomace or peels can achieve somewhat higher concentrations, but overall yields remain modest compared with the demand for nutraceutical?grade lycopene. Therefore, the development of alternative process including microbial based production of lycopene are being explored. Escherichia coli stands out as a preferred host due to its rapid growth, well-characterized genetics, and amenability to engineering. It can grow on cheap carbon sources such as glucose, glycerol, or agricultural waste, which lowers production costs for bio?based chemicals and natural products. In this study, we engineered E. coli for whole-cell biocatalytic lycopene production using a two-plasmid system. We co-expressed enzymes from the mevalonate pathway to boost isoprenoid precursors, alongside lycopene biosynthetic genes (e.g., crtE, crtB, crtI) that channel these intermediates into lycopene. This platform converts glucose directly into lycopene through multi-step biosynthesis, bypassing extraction bottlenecks. Our approach highlights E. coli's potential as an efficient, sustainable biocatalyst for industrial carotenoid production.
| Published in | Advances in Bioscience and Bioengineering (Volume 14, Issue 1) |
| DOI | 10.11648/j.abb.20261401.12 |
| Page(s) | 7-16 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Synthetic Biology, Whole-Cell Biocatalysis, Lycopene, Mevalonate Biosynthetic Pathway, Isoprenoid Biosynthesis
Strain/Plasmid | Description | Reference |
|---|---|---|
MG1655 (DE3) | ΔendA ΔrecA (λ DE3) | - |
DH5α | fhuA2 Δ(argF-lacZ)U169 phoA glnV44 Φ80 Δ(lacZ)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17 | - |
pJBEI-6409 | p15A, CmR, PlacUV5, atoB, HMGS, HMGR, PlacUV5, mvk, PMK, PMD, idi, Ptrc, trGPPS, ls | [24] |
pJBEI-6409 ΔtrGPPS ΔLS | p15A, CmR, PlacUV5, atoB, HMGS, HMGR, PlacUV5, mvk, PMK, PMD, idi | This study |
p5T7-LYCipi-ggpps | pSC101, SpR, PT7lacUV, ggpps, ipi, crtI, crtB | [25] |
Genes | Origin (Accession Number) |
|---|---|
ggpps | Taxus canadensis (AAD16018.1), codon optimized, truncated first 98 amino acids, methionine added |
crtI | AFZ89042.1 |
crtB | AFZ89043.1 |
ipi | AAA64978.1 |
LS | Mentha spicata (AAC37366.1), codon optimized |
gpps | Abies grandis (AAN01134.1), codon optimized |
ispA | E. coli (WP_097750737.1) |
idi | E. coli (AAD26812.1) |
atoB | E. coli (NC_000913.3) |
HMGS | Staphylococcus aureus |
HMGR | Staphylococcus aureus |
MK | Saccharomyces cerevisiae |
PMK | Saccharomyces cerevisiae |
PMD | Saccharomyces cerevisiae |
DMAPP | Dimethylallyl diphosphate (dimethylallyl pyrophosphate) |
GGPP | Geranylgeranyl Diphosphate (Geranylgeranyl Pyrophosphate) |
GGPPS | Geranylgeranyl Diphosphate Synthase |
IPP | Isopentenyl Diphosphate (Isopentenyl Pyrophosphate) |
MVA | Mevalonate Pathway |
MEP | 2‑C‑methyl‑d‑erythritol‑4‑phosphate pathway |
MEV | Mevalonate (MVA) Pathway Module in Plasmid |
T7 | T7 Bacteriophage Promoter System |
PlacUV5 | Hybrid lac‑UV5 promoter |
Ptrc | T7‑lac hybrid promoter |
IPTG | Isopropyl β‑D‑1‑thiogalactopyranoside |
OD600 | Optical density at 600 nm |
LB | Luria–Bertani (Luria–Broth) medium |
MG1655 | Escherichia coli K‑12 wild‑type strain MG1655 |
DH5α | E. coli cloning strain DH5α |
pJBEI‑6409 | p15A‑based plasmid encoding full MVA and downstream terpene genes |
pJBEI‑6409 ΔtrGPPS ΔLS | Engineered variant of pJBEI‑6409 lacking truncated GPPS and limonene synthase |
p5T7‑LYCipi‑ggpps | pSC101‑based T7‑promoter plasmid encoding lycopene pathway genes (ipi, crtI, crtB, ggpps) |
GGPPS | Geranylgeranyl Diphosphate Synthase (from Taxus canadensis) |
crtB | Phytoene Synthase (from Pantoea agglomerans) |
crtI | Phytoene Desaturase / lycopene synthase (from Pantoea agglomerans) |
ipi | Isopentenyl‑diphosphate isomerase (idi‑like gene from Pantoea agglomerans) |
trGPPS | Truncated Geranyl Diphosphate Synthase |
LS | Limonene Synthase |
MVK / MK | Mevalonate Kinase |
PMK | Phosphomevalonate Kinase |
PMD | Pyrophosphomevalonate Decarboxylase |
HMGS | HMG‑CoA Synthase |
HMGR | HMG‑CoA Reductase |
atoB | Acetyl‑CoA Thiolase (from E. coli) |
idi | Isopentenyl‑diphosphate Isomerase (native E. coli gene) |
| [1] | G. A. Armstrong and J. E. Hearst, “Genetics and molecular biology of carotenoid pigment biosynthesis,” FASEB J., vol. 10, no. 2, pp. 228-237, Feb. 1996, |
| [2] | Y. Xu et al., “Red and blue light promote tomato fruit coloration through modulation of hormone homeostasis and pigment accumulation,” Postharvest Biol. Technol., vol. 207, p. 112588, Jan. 2024, |
| [3] | V. M. Ye and S. K. Bhatia, “Pathway engineering strategies for production of beneficial carotenoids in microbial hosts,” Biotechnol. Lett., vol. 34, no. 8, pp. 1405-1414, Aug. 2012, |
| [4] | C. Wang et al., “Challenges and tackles in metabolic engineering for microbial production of carotenoids,” Microb. Cell Factories, vol. 18, no. 1, p. 55, Dec. 2019, |
| [5] | E. Giovannucci, “A Review of Epidemiologic Studies of Tomatoes, Lycopene, and Prostate Cancer,” Exp. Biol. Med., vol. 227, no. 10, pp. 852-859, Nov. 2002, |
| [6] | L. Bignotto et al., “Anti-inflammatory effect of lycopene on carrageenan-induced paw oedema and hepatic ischaemia-reperfusion in the rat,” Br. J. Nutr., vol. 102, no. 1, pp. 126-133, Feb. 2009, |
| [7] | J. W. Erdman, N. A. Ford, and B. L. Lindshield, “Are the health attributes of lycopene related to its antioxidant function?,” Arch. Biochem. Biophys., vol. 483, no. 2, pp. 229-235, Mar. 2009, |
| [8] | U. M. Khan et al., “Lycopene: Food Sources, Biological Activities, and Human Health Benefits,” Oxid. Med. Cell. Longev., vol. 2021, no. 1, p. 2713511, Jan. 2021, |
| [9] | R. Ciriminna, A. Fidalgo, F. Meneguzzo, L. M. Ilharco, and M. Pagliaro, “Lycopene: Emerging Production Methods and Applications of a Valued Carotenoid,” ACS Sustain. Chem. Eng., vol. 4, no. 3, pp. 643-650, Mar. 2016, |
| [10] | K.-W. Kong, H.-E. Khoo, K. N. Prasad, A. Ismail, C.-P. Tan, and N. F. Rajab, “Revealing the Power of the Natural Red Pigment Lycopene,” Molecules, vol. 15, no. 2, pp. 959-987, Feb. 2010, |
| [11] | T. Ma, Z. Deng, and T. Liu, “Microbial production strategies and applications of lycopene and other terpenoids,” World J. Microbiol. Biotechnol., vol. 32, no. 1, p. 15, Jan. 2016, |
| [12] | S. Yoon et al., “Enhanced lycopene production in Escherichia coli engineered to synthesize isopentenyl diphosphate and dimethylallyl diphosphate from mevalonate,” Biotechnol. Bioeng., vol. 94, no. 6, pp. 1025-1032, Aug. 2006, |
| [13] | M. Rohmer, M. Knani, P. Simonin, B. Sutter, and H. Sahm, “Isoprenoid biosynthesis in bacteria: a novel pathway for the early steps leading to isopentenyl diphosphate,” Biochem. J., vol. 295, no. 2, pp. 517-524, Oct. 1993, |
| [14] | J. Yang et al., “Bio-isoprene production using exogenous MVA pathway and isoprene synthase in Escherichia coli,” Bioresour. Technol., vol. 104, pp. 642-647, Jan. 2012, |
| [15] | Y. Boucher and W. F. Doolittle, “The role of lateral gene transfer in the evolution of isoprenoid biosynthesis pathways,” Mol. Microbiol., vol. 37, no. 4, pp. 703-716, Aug. 2000, |
| [16] | M. Li, R. Nian, M. Xian, and H. Zhang, “Metabolic engineering for the production of isoprene and isopentenol by Escherichia coli,” Appl. Microbiol. Biotechnol., vol. 102, no. 18, pp. 7725-7738, Sep. 2018, |
| [17] | W. R. Farmer and J. C. Liao, “Precursor Balancing for Metabolic Engineering of Lycopene Production in Escherichia coli,” Biotechnol. Prog., vol. 17, no. 1, pp. 57-61, Feb. 2001, |
| [18] | B. Du, M. Sun, W. Hui, C. Xie, and X. Xu, “Recent Advances on Key Enzymes of Microbial Origin in the Lycopene Biosynthesis Pathway,” J. Agric. Food Chem., vol. 71, no. 35, pp. 12927-12942, Sep. 2023, |
| [19] | C. Schwartz, K. Frogue, J. Misa, and I. Wheeldon, “Host and Pathway Engineering for Enhanced Lycopene Biosynthesis in Yarrowia lipolytica,” Front. Microbiol., vol. 8, p. 2233, Nov. 2017, |
| [20] | P. K. Ajikumar, K. Tyo, S. Carlsen, O. Mucha, T. H. Phon, and G. Stephanopoulos, “Terpenoids: Opportunities for Biosynthesis of Natural Product Drugs Using Engineered Microorganisms,” Mol. Pharm., vol. 5, no. 2, pp. 167-190, Apr. 2008, |
| [21] | G. Stephanopoulos, H. Alper, and J. Moxley, “Exploiting biological complexity for strain improvement through systems biology,” Nat. Biotechnol., vol. 22, no. 10, pp. 1261-1267, Oct. 2004, |
| [22] | P. Moroz et al., “Advances in Lycopene Production: From Natural Sources to Microbial Synthesis Using Yarrowia lipolytica,” Molecules, vol. 30, no. 21, p. 4321, Nov. 2025, |
| [23] | Y. Zhou, Y. Yao, F. Zhang, N. Yu, B. Wang, and B. Tian, “Enhancement of Lycopene Biosynthesis Using Self-Assembled Multi-Enzymic Protein Cages,” Microorganisms, vol. 13, no. 4, p. 747, Mar. 2025, |
| [24] | J. Alonso-Gutierrez et al., “Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production,” Metab. Eng., vol. 19, pp. 33-41, Sep. 2013, |
| [25] | A. O. Chatzivasileiou, V. Ward, S. M. Edgar, and G. Stephanopoulos, “Two-step pathway for isoprenoid synthesis,” Proc. Natl. Acad. Sci., vol. 116, no. 2, pp. 506-511, Jan. 2019, |
| [26] | J. Sambrook and D. W. Russell, “Preparation and Transformation of Competent E. coli Using Calcium Chloride,” CSH Protoc., vol. 2006, no. 1, p. pdb.prot3932, Jun. 2006, |
| [27] | V. J. J. Martin, D. J. Pitera, S. T. Withers, J. D. Newman, and J. D. Keasling, “Engineering a mevalonate pathway in Escherichia coli for production of terpenoids,” Nat. Biotechnol., vol. 21, no. 7, pp. 796-802, Jul. 2003, |
APA Style
Prajapati, G., Srivastava, K. R. (2026). Development of a Whole-Cell Biocatalytic System for Lycopene Production in Escherichia coli. Advances in Bioscience and Bioengineering, 14(1), 7-16. https://doi.org/10.11648/j.abb.20261401.12
ACS Style
Prajapati, G.; Srivastava, K. R. Development of a Whole-Cell Biocatalytic System for Lycopene Production in Escherichia coli. Adv. BioSci. Bioeng. 2026, 14(1), 7-16. doi: 10.11648/j.abb.20261401.12
@article{10.11648/j.abb.20261401.12,
author = {Gaurav Prajapati and Kinshuk Raj Srivastava},
title = {Development of a Whole-Cell Biocatalytic System for Lycopene Production in Escherichia coli},
journal = {Advances in Bioscience and Bioengineering},
volume = {14},
number = {1},
pages = {7-16},
doi = {10.11648/j.abb.20261401.12},
url = {https://doi.org/10.11648/j.abb.20261401.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.abb.20261401.12},
abstract = {Lycopene is one of the most well-known carotenoids in nature due to its high antioxidant properties and wide health benefits. Due to a wide range of applications in food, nutraceuticals, cosmetics, and pharmaceuticals, demand for lycopene has grown significantly and its unmet need requires production at large scale. Conventionally, lycopene extraction from natural sources suffers from extensive downstream processing and poor yield. Industrial processes using tomato pomace or peels can achieve somewhat higher concentrations, but overall yields remain modest compared with the demand for nutraceutical?grade lycopene. Therefore, the development of alternative process including microbial based production of lycopene are being explored. Escherichia coli stands out as a preferred host due to its rapid growth, well-characterized genetics, and amenability to engineering. It can grow on cheap carbon sources such as glucose, glycerol, or agricultural waste, which lowers production costs for bio?based chemicals and natural products. In this study, we engineered E. coli for whole-cell biocatalytic lycopene production using a two-plasmid system. We co-expressed enzymes from the mevalonate pathway to boost isoprenoid precursors, alongside lycopene biosynthetic genes (e.g., crtE, crtB, crtI) that channel these intermediates into lycopene. This platform converts glucose directly into lycopene through multi-step biosynthesis, bypassing extraction bottlenecks. Our approach highlights E. coli's potential as an efficient, sustainable biocatalyst for industrial carotenoid production.},
year = {2026}
}
TY - JOUR T1 - Development of a Whole-Cell Biocatalytic System for Lycopene Production in Escherichia coli AU - Gaurav Prajapati AU - Kinshuk Raj Srivastava Y1 - 2026/04/02 PY - 2026 N1 - https://doi.org/10.11648/j.abb.20261401.12 DO - 10.11648/j.abb.20261401.12 T2 - Advances in Bioscience and Bioengineering JF - Advances in Bioscience and Bioengineering JO - Advances in Bioscience and Bioengineering SP - 7 EP - 16 PB - Science Publishing Group SN - 2330-4162 UR - https://doi.org/10.11648/j.abb.20261401.12 AB - Lycopene is one of the most well-known carotenoids in nature due to its high antioxidant properties and wide health benefits. Due to a wide range of applications in food, nutraceuticals, cosmetics, and pharmaceuticals, demand for lycopene has grown significantly and its unmet need requires production at large scale. Conventionally, lycopene extraction from natural sources suffers from extensive downstream processing and poor yield. Industrial processes using tomato pomace or peels can achieve somewhat higher concentrations, but overall yields remain modest compared with the demand for nutraceutical?grade lycopene. Therefore, the development of alternative process including microbial based production of lycopene are being explored. Escherichia coli stands out as a preferred host due to its rapid growth, well-characterized genetics, and amenability to engineering. It can grow on cheap carbon sources such as glucose, glycerol, or agricultural waste, which lowers production costs for bio?based chemicals and natural products. In this study, we engineered E. coli for whole-cell biocatalytic lycopene production using a two-plasmid system. We co-expressed enzymes from the mevalonate pathway to boost isoprenoid precursors, alongside lycopene biosynthetic genes (e.g., crtE, crtB, crtI) that channel these intermediates into lycopene. This platform converts glucose directly into lycopene through multi-step biosynthesis, bypassing extraction bottlenecks. Our approach highlights E. coli's potential as an efficient, sustainable biocatalyst for industrial carotenoid production. VL - 14 IS - 1 ER -
Medicinal and Process Chemistry Division, CSIR-Central Drug Research Institute, Lucknow, India;Academy of Scientific and Innovative Research (AcSIR), Ghaziabad, India
Medicinal and Process Chemistry Division, CSIR-Central Drug Research Institute, Lucknow, India;Academy of Scientific and Innovative Research (AcSIR), Ghaziabad, India
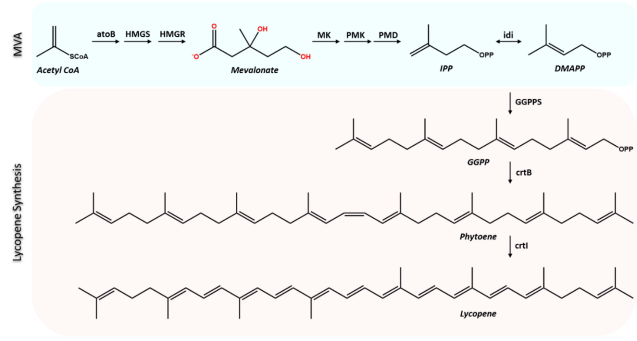
Figure 1. Schematic representation of lycopene biosynthetic route.
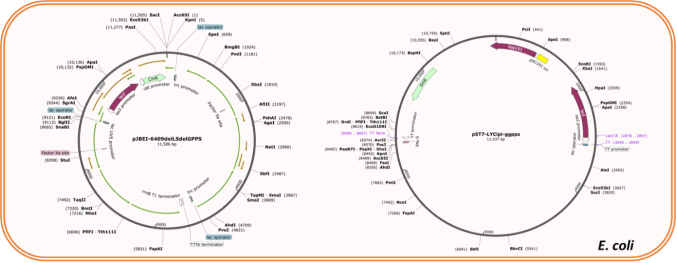
Figure 2. Co-transformation strategy showing pJBEI-6409 ΔtrGPPS ΔLS and p5T7-LYCipi-ggpps in E. coli.
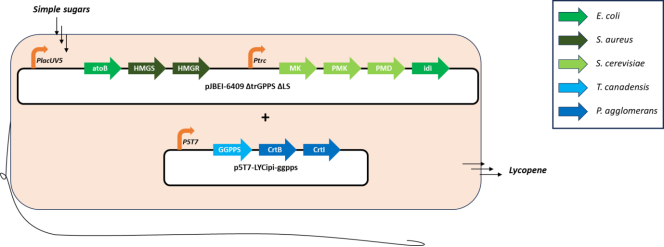
Figure 3. The heterologous mevalonate (MVA) pathway and lycopene biosynthetic pathway introduced into E. coli to produce lycopene. The MVA pathway utilises acetyl-CoA derived from glucose to produce isoprenoid metabolites. IPP, Isopentenylpyrophosphate; DMAPP, Dimethylallypyrophosphate; GGPP, Geranylgeranylpyrophosphate.

Figure 4. a) The red pigmentation confirms the production of lycopene in E. coli MG1655 incorporating plasmids pJBEI-6409 ΔtrGPPS ΔLS and p5T7-LYCipi-ggpps(right) with respect to control E. coli MG1655 carrying plasmid p5T7-LYCipi-ggpps (middle) and E. coli MG1655 (left) devoid of both plasmids. b) Culture after 120 hours of E. coli MG1655 incorporating plasmids pJBEI-6409 ΔtrGPPS ΔLS and p5T7-LYCipi-ggpps(middle) with respect to control E. coli MG1655 carrying plasmid p5T7-LYCipi-ggpps (right) and E. coli MG1655 (left) devoid of both plasmids. c) Cell pellet of the whole-cell culture containing lycopene metabolite. d) Pilot scale of 4 L lycopene culture in a 10 L scale fermenter unit present in CSIR-CDRI facility. e) Collected 4 L lycopene culture in two 2 L flask.
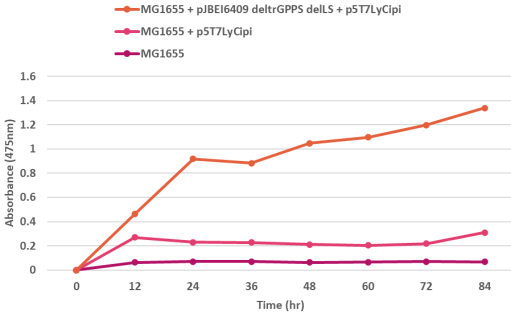
Figure 5. Lycopene production assessed by UV/Vis spectrophotometer at 475 nm at different time intervals. Absorption of lycopene in E. coli MG1655 strain harbouring pJBEI6409 ΔtrGPPS ΔLS and p5T7-LYCipi-ggpps was recorded between 1.2-1.4 as compared to E. coli MG1655 strain harbouring only p5T7-LYCipi-ggpps and E. coli MG1655 strain between 0.2-0.4 and 0-0.2 upto 84 hours.

